Serious writing for
serious readers
Summary
6 December 2010
This is a short book, only 120 pages excluding endnotes, published in 2008. I found it impossible to put down. As well as the specific topics covered, it provides an excellent simple introduction to the way that science is enabling us to understand the history of populations.
We each get our genetic inheritance from our parents. For most of our chromosomes this involves mixing the inheritance from our father and our mother. However only males have a Y chromosome, and this comes entirely from our father with no genetic input from our mother.
Accordingly, if there were no mutations (changes to the coding of the DNA) my Y chromosome would be identical to the Y chromosome of every one of my paternal ancestors from the dawn of time, and more importantly would be identical to the Y chromosome of every living man descended from my earliest paternal ancestor.
However, while most of the time the Y chromosome is inherited perfectly from the father without any change, occasionally there is a mutation. Once that mutation arises, the mutated Y chromosome is passed down the male generations. That is why there are small differences in the Y chromosome between most men unless they are closely related with a common male ancestor who is not too far back in the family tree. The further back in the family tree you go, the greater the likelihood of mutations having arisen during the intervening generations. Furthermore, while most of the Y chromosome suffers relatively few mutations, parts known as microsatellites tend to mutate more rapidly.
Every cell in our body contains a nucleus which is where our 23 pairs of chromosomes can be found. In addition, each cell contains mitochondria which are like small self-contained organisms swimming around in the fluid between the cell nucleus and the cell wall. These mitochondria also contain DNA, although far less DNA than the chromosomes found in the cell nucleus. The key point about mitochondrial DNA is that it is inherited entirely from your mother with no involvement from your father's genetic inheritance.
Mitochondrial DNA also mutates and it provides a way of tracing maternal ancestry in the same way that paternal ancestry is traced through the Y chromosome. The above points are illustrated in diagram below:
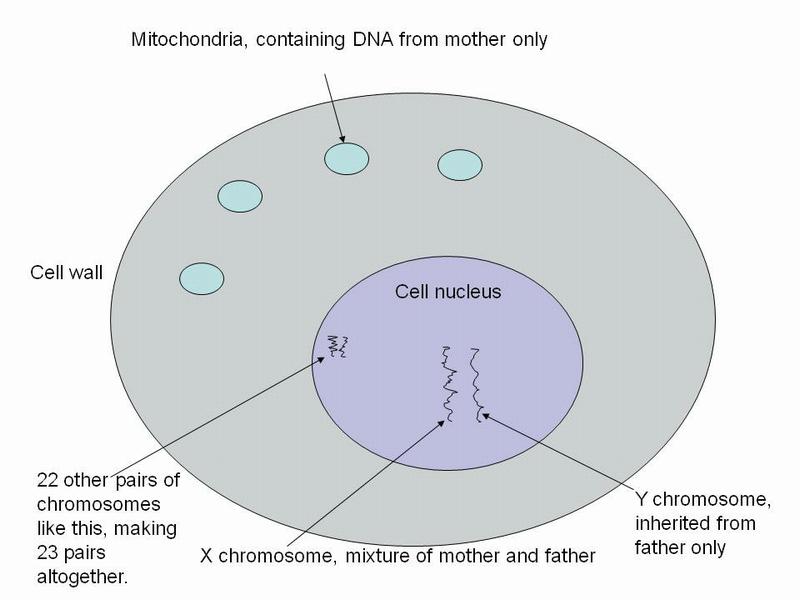
Diagram of a cell showing the nucleus and its chromosomes and the mitochondria
23andMe is a commercial website which sells services as well as providing free information about genetics. As far as I am aware, neither I nor any member of my family has any connection with it.
If you wish to learn more about genetics, I suggest reading the genetics 101 pages at the website 23andMe.
The book looks at five main topics and these are so interesting that they are worth reviewing individually.
Chapter 3 of the book of Numbers explains that Aaron's male descendants were anointed as priests. The word priest in Hebrew is Cohen, plural Cohanim. The Cohanim were responsible for conducting animal sacrifices in the Temple in Jerusalem.
After the Temple was destroyed by the Romans in 70 A.D., the Cohanim had no further religious function and religious leadership in the Jewish community passed to the rabbis. However many Jewish men regard themselves as being descended from the Cohanim and will bless their fellow Jews at the Western Wall and in synagogues on the high holy days. As an aside, the hand gesture used for this blessing is the same as that used by Mr Spock in Star Trek as it was introduced into the series by Leonard Nimoy who is Jewish.
The author set out to address whether this belief in descent from the Cohanim has any validity. To quote:
We can imagine two very different sorts of answers; adoption and genetic continuity. The Jews have long carried with them the story of the priests, and at some point in their history the stories could have motivated people to assume, or adopt, priestly status, regardless of their genetic ancestry. In other words, sometime after the dispersal in the first and second centuries CE, a group of Jews (or may be non-Jews, a possibility I consider in chapter 3) could have decided to adopt the title of priest or been awarded it. In time, this group could have come to be accepted as such. We know that in the Second Temple period the award of priestly status had everything to do with political expediency and relatively little to do with genealogy.
At the other extreme from this nongenetic adoption of status is the possibility of genetic continuity, which assumes that the oral tradition is largely correct; the Cohanim of today are indeed culturally and genetically continuous (on the male side) with the priests who survived the destruction of the second Temple and subsequent uprisings.
The author and his team first decided to ascertain what proportion of Jews today might be considered as being Cohanim. They addressed this by going into Jewish cemeteries and counting the priestly hand symbol on the tombstones, since only the tombstones of men classifying themselves as Cohanim have this symbol. From this, they estimated that 4-5% of male Jews are Cohanim. That would mean about 500,000 men worldwide and if the tradition were accurate, all of them would have a Y chromosome derived from Aaron about 3,000 years ago.
If the oral tradition were true, then the Y chromosomes of the Cohanim would be different from the general Jewish population in a particular way; “they be much less heterogeneous. In the general Jewish population, we would expect many types of Y chromosomes; those of Jews and convert to Judaism from all over, representing all different occupations and genetic origins.”
The team collected mouth swabs in Canada, Israel and the UK ensuring that the individuals swabbed did not have common grandparents. Swabs were taken from Jews in general and from Jews self identifying as Cohanim. The team preferred self identification to relying on surnames such as Cohen, Coen, Kagen, Cohn etc. The swab donors were also asked whether they were Ashkenazi or non-Ashkenazi.
With a total sample size of 306 men, the team using both mutations and microsatellites identified 109 different types of Y chromosome. Despite the high level of overall variation, they found that the most common chromosomes amongst non-Cohanim were found in only 12% of the sample. Conversely more than half of the Cohen Y chromosomes were identical at all the sites considered, and this particular type of Y chromosome was found at high frequencies in both Ashkenazi and Sephardi Cohanim. They called this chromosomal type the Cohen Modal Haplotype. Modal means most frequent and haplotype indicates a particular collection of variant forms. Furthermore, they found that amongst the Cohanim many of the other types of Y chromosome were very similar to the Cohen Modal Haplotype. This cluster of chromosomes accounts for 64% of the sample amongst the Cohanim but only 14% of the sample amongst non-Cohanim.
The overall conclusion is that both Ashkenazi and Sephardi Cohanim share a common ancestor. That common ancestor must have lived before the time when Jews split into Ashkenazi and Sephardi communities. Furthermore, because there is a cluster of related types of Y chromosome, it is possible to estimate how far back they started to diverge and this appears to be about 1,000 BC which roughly corresponds to the time of Solomon's Temple.
The science cannot prove that Cohanim are descended from Aaron. However it does show that, unlike Jews in general, most Cohanim are descended from a single common male ancestor who lived about 1,000 BC.
In South Africa is a Bantu speaking tribe called the Lemba. There are about 70,000 of them and they claim that they arrived in South Africa from Judaea via a place called Sena.
The Lemba have a number of interesting characteristics which are similar to Jewish practices. They are monotheists and do not practice idol worship. They do not take non-Lemba spouses. They practice circumcision. Lemba are buried horizontally whereas most other African tribes bury their dead in a sitting position. Lemba practice animal sacrifice using special knives with prayers of thanks. The Lemba do not eat pork and also do not mix milk and meat.
The author and his team collected genetic samples from the Lemba, from local non-Lemba Bantu speakers in South Africa and from the Yemen since the Lemba claim to have passed through the Yemen on their way to South Africa.
The Y chromosomes of non-Cohanim Jews were very similar to the Y chromosomes of Yemenis but both were very different from the non-Lemba South African Bantu speakers. This was to be expected since both the Jews and Yemenis are Semites and different from South African Bantu speakers. Interestingly, one key point is that the virtually no Yemenis had the Cohen Modal Haplotype (CMH) discussed above even though it is common amongst Jews. Out of 100 Yemenis, only a single case of the CMH was observed and none amongst the South African Bantu speakers.
Because the Y chromosomes of Bantu speakers and Semites differ, the team was able to go through the Lemba Y chromosomes one by one and classify them as either Bantu or Semitic in origin. They found that two thirds of Lemba Y chromosomes have a Semitic source rather than a Bantu speaking source. Even more significantly, nearly 10% of Lemba Y chromosomes are in the CMH category which gives a specifically Jewish source rather than merely Semitic.
Furthermore, although the Semitic Y chromosomes were widespread through the Lemba clans, most of the CMH chromosomes were observed in only one Lemba clan, the Buba. The CMH was found at frequencies over 50% in Buba from distinct geographic areas. The Lemba oral traditions regard the Buba lineage as going back to the leadership that led them from Judaea into Yemen.
The reasonable overall conclusion is that, as far as the male line is concerned, the genetic data supports the Lemba narrative of descent from Jewish people.
The Cohanim mentioned above were a subset of the tribe of Levi, being the descendants of Aaron. However the entire tribe of Levi was regarded as sacred although as a sort of "second-class priesthood." For example the Cohanim had first chance to read from the Torah while the Levites were to read second. The Cohanim conducted temple sacrifices whereas Levites had more mundane Temple tasks such as cleaning, guarding the door and transporting the Ark of the Covenant.
Today in synagogues Levites are called second after Cohanim to read passages from the Torah and in some congregations they still wash the hands of the Cohen before the Cohen delivers his blessing.
The author uses the term Israelites to refer to Jews who are not Cohanim and are not Levites. The Cohanim study mentioned above found that present-day Ashkenazi and Sephardi Cohanim are more genetically similar to one another than they are to either Israelites or non-Jews. However Levites turned out to have as many different Y-chromosome types as Israelites generally. Intriguingly, the Levites chromosomes were not a subset of the Israelites. Ashkenazi Levites have a very high frequency of a particular collection of Y chromosome markers (a haplogroup) called R-M17 which is rare in Jewish populations and generally rare or absent altogether in populations of the near East. Accordingly the question is where this haplogroup R-M17 came from.
The patterns of the R-M17 Y chromosomes did not resemble general Jewish Y chromosomes implying that they were unlikely to have come from the ancestral Hebrew population. Instead they appear to have a foreign origin but had found their way into Ashkenazi Levites but not into other Jewish groups.
In the past, European Jews moved eastwards from Germany into Poland, Lithuania and Russia. Accordingly for many centuries they lived between German-speaking peoples to the West and Slavic speakers further east. The author’s team collected genetic samples from the non-Jewish populations of northern Germany and Belarus. As well as Slavic speaking Belarus, the team collected samples from Sorbs (not to be confused with Serbs) who are a Slavic speaking minority of about 60,000 people in Germany concentrated in the eastern corner near the Czech Republic.
As the control group, the team also collected samples from another population that had some R-M17 Y chromosomes but which should not be a source for the Levites’ Y chromosomes. They chose the population of Norway since Norway prohibited all Jewish immigration throughout the historical period under consideration, only permitting Jews to settle in there since 1851.
Amongst the Israelites, less than 5% of the Y chromosomes contained R-M17. Amongst Belarusians and Sorbs the frequencies were 50% and 66% respectively. Amongst the Levites, 56% of the Y chromosomes were R-M17, very similar to the Slavic groups. Germans and Norwegians R-M17 frequencies were 13% and 22%, much lower than the Slavs but much higher than the Israelites. The Sorbs and the Ashkenazi Levites were statistically indistinguishable.
The above findings do not require the Sorbs to be the source of the Levite Y chromosomes; both could have had a different common source.
The team then looked at more detailed microsatellite variations. To quote:
Using more extensive microsatellite variation, we found that among the Levites the Y chromosomes were mostly close relatives of one another. They formed a tight group of just three haplogroups, separated from one another by single microsatellite mutations. Furthermore, the three most common types were either completely absent or else barely present in the Sorbs and Belarusians. The case for Slavic origins of Ashkenazi Levites was showing some serious cracks.
At this stage, the data has already shown that whereas Cohanim status was largely transmitted by male inheritance, the same is not true of Levite status. However, that still leaves the question of where the Levite R-M17 came from.
The author does not reach a conclusion. However he speculates that the Khazars could be the source. The Khazars had an empire in Central Asia along the Caspian Sea and converted to Judaism in the ninth century. In his book, "The Thirteenth Tribe” published in 1976, Arthur Koestler speculated that Ashkenazi Jewry may have a greater genetic inheritance from the Khazars than from the ancient Israelites. More work is needed. In particular the author points out that the modern language most closely related to that of the Khazars is Chuvash which is spoken by more than 1 million citizens of the Chuvash Republic within the Russian Federation. Examination of Chuvash Y chromosomes would appear to be the next step.
However, at present the origin of the non-Jewish elements in the Levite Y chromosome remains an unresolved mystery.
In almost all cultures, including ancient Israel while it occupied the Holy Land, legitimacy and descent follow the male lineage. To quote from the book:
We have no indication that the mother determined ethnic or religious identity in the ancient near East in general or amongst the ancient Israelites in particular. Indeed, there are many indications to the contrary. In the Bible, people are often named after male founders: Moabites after Moab, Ammonites after Ben-Ammi, and Edomites after Jacob's brother Esau. More directly, when Old Testament parents have different backgrounds, the children are usually identified as members of the father's ethnic group. Ruth, for example, is a Moabite who marries an Israelite of the Ephrathite clan, but when the Bible introduces her famous great-grandson, it says "David was the son of an Ephrathite named Jesse."
The author states that it is unclear when the diaspora practice of tracing Jewish lineage through the mother developed but it is clear that by 550AD-700AD it was fully established.
As mentioned above, mitochondrial DNA is inherited only from the mother. The author and his team set out to review paired comparisons of Jewish and host populations with the intention of determining how much of the given Jewish population’s gene pool was derived from an ancestral Israelite population and how much from the local host population via religious conversion or other means of gene flow.
The team chose 18 populations from Europe, North Africa, the Middle East, Central Asia and the Indian subcontinent, in each case comparing the Jewish community with a neighbouring non-Jewish population. They also looked at a set of Arabs from Israel and Palestine to represent possible long-term residents of the biblical land of Israel.
The working hypothesis was that female defined ethnicity would block or slow down gene flow from host populations into Jewish populations along the female line but not along the male line. If the flow rates of mitochondrial DNA and Y chromosomes were significantly different, with the mitochondrial DNA flow rate being much lower, this would confirm that the Jewish practice of descent down the female line had indeed been followed. Unfortunately, the results were inconclusive.
After deliberation, the team decided to look at the raw data on mitochondrial DNA variation in the individual Jewish populations themselves. The results were striking. Amongst Georgian Jews, more than half of the individuals had a single mitochondrial DNA type which was hardly found anywhere else in the world. Moroccan Jews showed a similar pattern but with a different mitochondrial DNA type and so did the Bene Israel (a Jewish population from Mumbai.) Conversely, the Y chromosomes showed hardly any differences between the Jews and their host populations in terms of genetic diversity.
A dramatic loss of genetic diversity normally arises from a dramatic shrinkage in the population or from a "founder event." An example of a founder event would be colonists in a new area all marrying a number of sisters as wives; all future offspring would have the mitochondrial DNA of the mother of those sisters.
This distribution of mitochondrial DNA found amongst the Jewish populations studied is unusual.
Geneticists find that in most human populations, variations in the Y chromosome increase with geographical distance whereas mitochondrial DNA remains much more similar over large geographical distances. The hypothesis for this general distribution is patrilocality; the tendency for men to remain in one place while women move from their place of birth to join their future husbands. For example this was the normal pattern in the Punjab villages in India where my own mother and father grew up. As a result of the men remaining in fixed locations, the Y chromosomes of different locations diverge over time due to mutation. Conversely variations in mitochondrial DNA would be smoothed out as women moved from location to location to join new family groups.
As noted above, with Jewish populations the pattern was the opposite. The mitochondrial DNA showed a much greater variation than the Y chromosomes, whereas the host populations from the same geographic areas showed the normal pattern of greater Y chromosome variation. Accordingly the Jewish pattern is not dependent upon the specific geographic areas the Jewish populations are located in; it is something specific to the Jewish groups themselves arising from their behaviour and marriage patterns.
The author speculates that Jewish men travelled long distances (for example along the Silk Road) to establish small Jewish communities. They would take local women for wives with the communities later being augmented by additional male travellers from Jewish source populations. However once the new communities were established barriers would go up against the further input of new mitochondrial DNA because of female defined ethnicity; non-Jewish women would not be permitted to join the group as wives or otherwise. This is hypothetical but it would explain the genetic data.
The author points out that there is no reason to believe that Jews suffer more from genetic disease than Africans, Arabs, Icelanders or any other population. However he states that a search of the database www.pubmed.com for the term "Ashkenazi" found more than 1300 papers. (2,702 when I performed the search on 6 December 2010!) The obvious explanation is ascertainment bias; Ashkenazi Jews are primarily urban, live near major clinical and research centres and have a long involvement with medical research.
As a genetic historian, the author wants to understand when so-called Jewish disease mutations originated. He states that at least 40 conditions determined by single genes have been described in Jewish populations and perhaps half of these occur at a higher frequency among Jews than among randomly chosen people.
There are two principal contending explanations why certain diseases have become entrenched amongst Jewish populations:
This results from a small population settling in a new area, causing the community that develops to be genetically different from the larger source population that it came from, depending upon the precise genetic make up of the small number of founders of the new group. Furthermore, Jewish populations have often suffered major persecutions resulting in sharp reductions in population size giving rise to so-called genetic bottlenecks which reduce future genetic diversity.
This occurs when carriers of a genetic disorder receive benefits from being carriers. The classic example is sickle cell anaemia, a recessive genetic disorder where carriers do not suffer from sickle cell disease, as they only have one copy of the defective gene, but are protected from malaria. Accordingly in certain parts of Africa the frequency of being a carrier for sickle cell disease can be as high as 50%.
The debate between these alternative explanations is very contentious. For example, Tay-Sachs disease is a recessive disorder which kills children in early life. Before the introduction of widespread screening, it was 100 times more frequent amongst Ashkenazi infants than amongst the general population. Amongst North American Jews, one person in every 31 carries a Tay-Sachs mutation. While a single mutation in the HEXA gene accounts for the majority of Tay-Sachs disease (73%), two other mutations explain another 20% with the remainder being due to rare mutations which nearly 80 have been described.
Supporters of the selection hypothesis point out that there are at least four distinct diseases, Tay-Sachs, Gaucher, Niemann-Pick and mucolipidosis IV which occur at increased frequencies in the Ashkenazi population. Each one is a lysosomal disorder and part of the same biochemical pathway involving the storage of specialised lipid molecules known as glycosphingolipids. The argument is that four mutations occurring at random in the same part of the genome is unlikely unless there is some selective advantage for carriers of these mutations; an argument strengthened by the fact that multiple mutations can be found in the case of each disease rather than a single primary mutation.
The author mentions a paper by three anthropologists, Cochran, Hardy and Harpending. This proposes that the demographic and sociological history of Ashkenazim in mediaeval Europe created an environment that made intelligence a highly advantageous trait. Accordingly certain gene variants that conferred intelligence were selected for and in particular carriers for mutations in lysosomal storage genes were likely to exhibit increased intelligence.
This argument is controversial and at present there is insufficient evidence to decide either way.
Our knowledge of the human genome is still relatively limited and we have only recently develop the technologies that allow us to sequence large amounts of DNA at a low cost. In the near future, far more people will have their DNA sequenced which will provide significant amounts of new data for genetic historians.
The pioneering work in genetic history outlined in this book is an illustration of what we might learn. The book itself is extremely well written and completely gripping. It does not require a scientific background to be able to read it and I would recommend it to everyone.
Advertisement
Follow @Mohammed_Amin

